



讓你的lncRNA擺脫miRNA的束縛(上)
 2021-09-29 13:42:18
2021-09-29 13:42:18
來源/作者:普拉特澤生物-醫學整體課題外包

非編碼RNA近幾年來的熱度雖然有所降低,但瘦死的駱駝依然能發文章。如果沒有一個新穎的靶蛋白砸到腦袋上,非編碼RNA的萬精油套路仍然還是廣大基層科研工作者沖擊高分的首選。對于非編碼RNA家族的成員,各位同學最熟悉的當然是lncRNA和miRNA兩兄弟了。這兩兄弟通過一種叫做ceRNA(competitive endogenous RNA)的聽著挺高端,其實挺簡單的機制,硬是在過去的十來年往江湖里灌了不少的農夫山泉。

然鵝,好景不長,miRNA這不爭氣的娃娃近幾年率先顯了疲態,單獨研究miRNA的文章分值區間一再走低,miRNA漸漸地就變成了一個傍著lncRNA這個老大哥出現的小老弟。雖然以lncRNA/miRNA/靶基因為主軸的套路依然適用于3-5分的區間,甚至在你的lncRNA足夠新穎的情況下依然有能力沖擊更高的分數,但是有夢想的lncRNA也開始逐漸往新的方向拓展。
那么這次就由野菜君通過幾篇典型文獻來給各位同學介紹4種有別于ceRNA機制的lncRNA的研究方向,如果你手頭正好有一個熱乎的lncRNA,又不再想走miRNA的水路,或許能從里面找到一些靈感。
(一)
LncRNA與組蛋白甲基化(histone methylation)
本節關鍵詞:PRC2復合物,EZH2,H3K27三甲基化修飾
本節相關文獻:Long noncoding RNA AFAP1-AS1 predicts a poor prognosis and regulates non–small cell lung cancer cell proliferation by epigenetically repressing p21 expression,Molecular Cancer ,IF:10.6
首先我們來看一種lncRNA在告別miRNA后最常見的路子——組蛋白甲基化。組蛋白甲基化是調控基因表達的重要方式,主要包括H3K27me3,H3K9me3,H3K4me3等等。我們以H3K27me3這種與lncRNA的相關報道最多的修飾方式為例,其含義是組蛋白Histone 3 (H3) 的第27位的賴氨酸 (K) 被三甲基化(me3)修飾,而帶有這種修飾的組蛋白結合到靶基因的啟動子區域,就會抑制靶基因的表達。細胞內主要負責H3K27me3修飾的是PRC2(Polycomb Repressive Complex 2)這個復合物。然后呢,這個復合物主要由EZH2, SUZ12, EED等幾個亞基組成,其中EZH2(Enhancer Of Zeste 2)又是直接具有甲基化轉移酶催化活性的最重要的亞基。近年來,發現PRC2以及EZH2在腫瘤中上調,通過抑制多種抑癌基因的表達,發揮促癌作用。為了方便閱讀,我們可以通過一張模式圖對這個機制先有一個大概的印象。

好了,介紹完背景,我們通過一篇典型文獻來看看針對我們的lncRNA,如何研究它調節組蛋白甲基化的機制。這篇文獻18年發表在molecular cancer上(Long noncoding RNA AFAP1-AS1 predicts a poor prognosis and regulates non–small cell lung cancer cell proliferation by epigenetically repressing p21 expression)。值得一提地是,這個雜志影響因子近幾年的影響因子在5-7的區間徘徊,但是2018年神奇地躥升到了10.6分。關于lncRNA對腫瘤基礎表型的功能驗證就不在此贅述了,我們直接看機制部分結果。
首先,作者通過測序和GO分析,發現在A549細胞中敲除AFAP1-AS1能夠引起300多個基因上調兩倍以上(Fig. 4a)以及這些基因與細胞增殖(Fig. 4b)等過程有關。當然,這一步也不是必須的,我們完全可以通過公共數據庫的挖掘(比如TCGA)找出在某腫瘤中與AFAP1-AS1的表達呈負相關的抑癌基因,或直接像(Fig. 4c)和(Fig. 4d)的結果那樣,選取一些經典的抑癌因子進行qPCR的驗證,同時這些抑癌因子又往往是有報道被EZH2抑制的。總之,這一步就是找出調控AFAP1-AS1引起的差異表達基因。
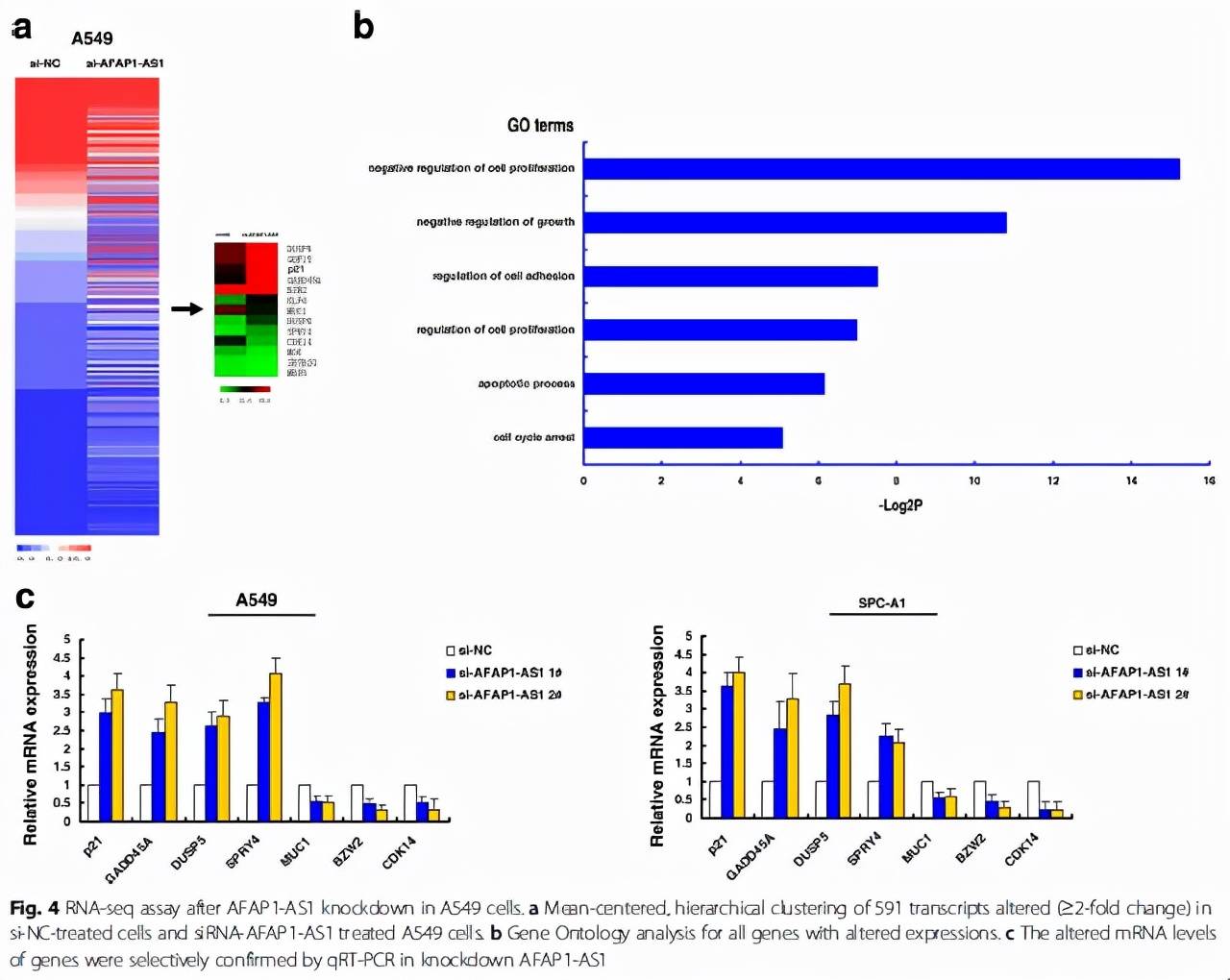
然后呢,作者先是驗證了AFAP1-AS1在核內和胞漿的表達分布,畢竟我們的H3K27me3修飾是發生在核內(Fig. 5a),再通過RIP的方式驗證了AFAP1-AS1與EZH2以及SUZ12均有結合(Fig. 5b)。
小拓展:對于具體lncRNA與EZH2結合后是通過何種方式發揮作用,這個機制大多數研究都是沒有再深究的。就連發表在PNAS上的lncRNA/EZH2套路文章也僅僅是探明了lncRNA上的一段富G的片段(G-rich motif)參與結合了EZH2,而關于EZH2上結合lncRNA的片段,可能存在于其N端的32–42 helix。有興趣的讀者可以自行參看這兩篇補充文獻深入了解(HERES, a lncRNA that regulates canonical and noncanonical Wnt signaling pathways via interaction with EZH2;Conserved RNA-binding specificity of polycomb repressive complex 2 is achieved by dispersed amino acid patches in EZH2) 。

最后,作者把下游的靶基因鎖定在p21這個經典的抑癌基因,進一步驗證了AFAP1-AS1和EZH2分別對p21表達的調控(Fig. 6a,Fig. 6b),同時,通過CHIP實驗,發現敲除AFAP1-AS1后,EZH2以及H3K27me3在p21啟動子區域的結合降低,最終導致p21的表達抑制被解除(Fig. 6c)。
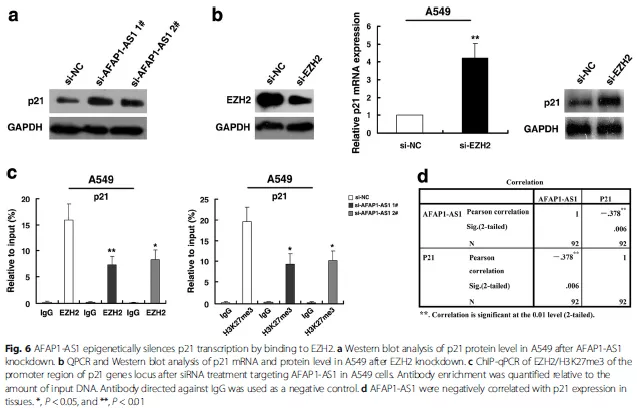
至此,作者用最低碳的方式,驗證了核內的AFAP1-AS1通過結合EZH2(RIP,RNA pull down實驗),將EZH2募集(recruitment)到靶基因p21的啟動子區域(CHIP實驗),EZH2通過將該區域的H3組蛋白進行H3K27me3修飾,抑制p21的表達(WB, qPCR驗證)。將之前的那個作用模式圖用具體的分子代替就是這樣:
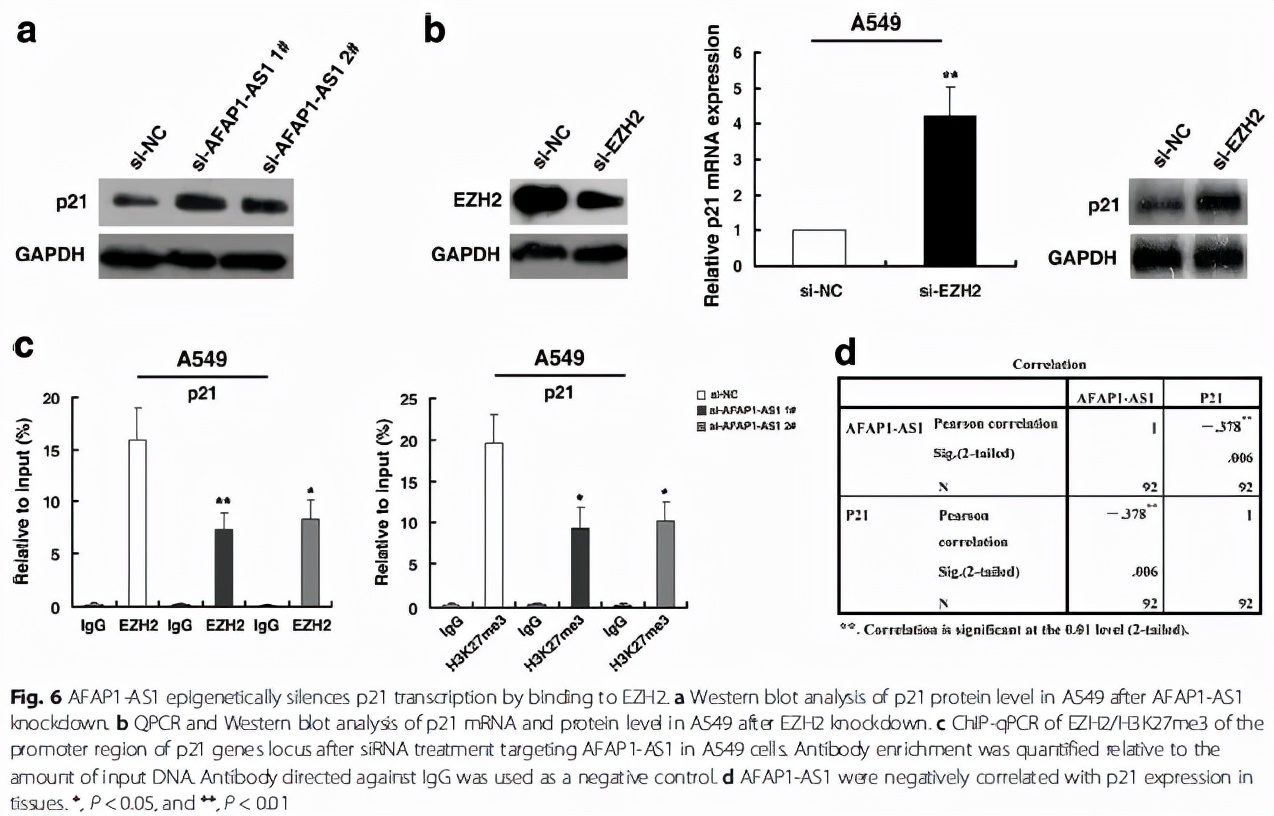
當然我們也可以參考其他一些lncRNA/EZH2/靶基因的文獻,進一步增加我們的實驗豐度,但是核心的(最低碳的)機制基本就是該文獻中展現的這樣。值得注意的是,一般來說,lncRNA并不會改變EZH2的整體表達,只是將EZH2招募到啟動子區域;而對于H3K27me3的整體表達,則存在lncRNA的差異性,有的會上調,有的則并不改變。
關于lncRNA與組蛋白甲基化的技術總結
1. 關鍵的驗證內容是lncRNA與EZH2的結合,以及lncRNA對EZH2, H3K27me3在靶基因啟動子區域的富集的調控。
2. 雖然此文獻中沒有展示,但這里強烈建議還是要加上RNA pull down的結果與RIP結果一道作為lncRNA結合EZH2的進一步的驗證。
3. 一般來說不需要再深入研究lncRNA與EZH2的作用機制。
4. 除了H3K27me3,還存在如H3K4me3,H3K9me3的修飾,如果你手頭的lncRNA已經有了與EZH2的報道,不妨嘗試其他幾種修飾。
(二)
LncRNA與RNA結合蛋白(RNA bind protein, RBP)
本節關鍵詞:mRNA穩定性
本節相關文獻:Long Noncoding RNA MEG3 Induces Cholestatic Liver Injury by Interaction With PTBP1 to Facilitate Shp mRNA Decay,Hepatology,IF:14
接下來我們再看另一種lncRNA通過非ceRNA的機制調控基因表達的方式。RNA結合蛋白(RBP),顧名思義是一類可以與RNA結合的蛋白。RBP可以通過與mRNA結合,調控mRNA的剪切(splicing),穩定性(stability),翻譯起始(translation initiation)等過程,從而在翻譯階段調控基因的表達。我們通過這篇2017年發表于Hepatology(14分)上的文章(Long Noncoding RNA MEG3 Induces Cholestatic Liver Injury by Interaction With PTBP1 to Facilitate Shp mRNA Decay),來了解一下經典的lncRNA+RBP的套路。同樣,我們先看一張機制圖來對這個機制有一個初步的理解。
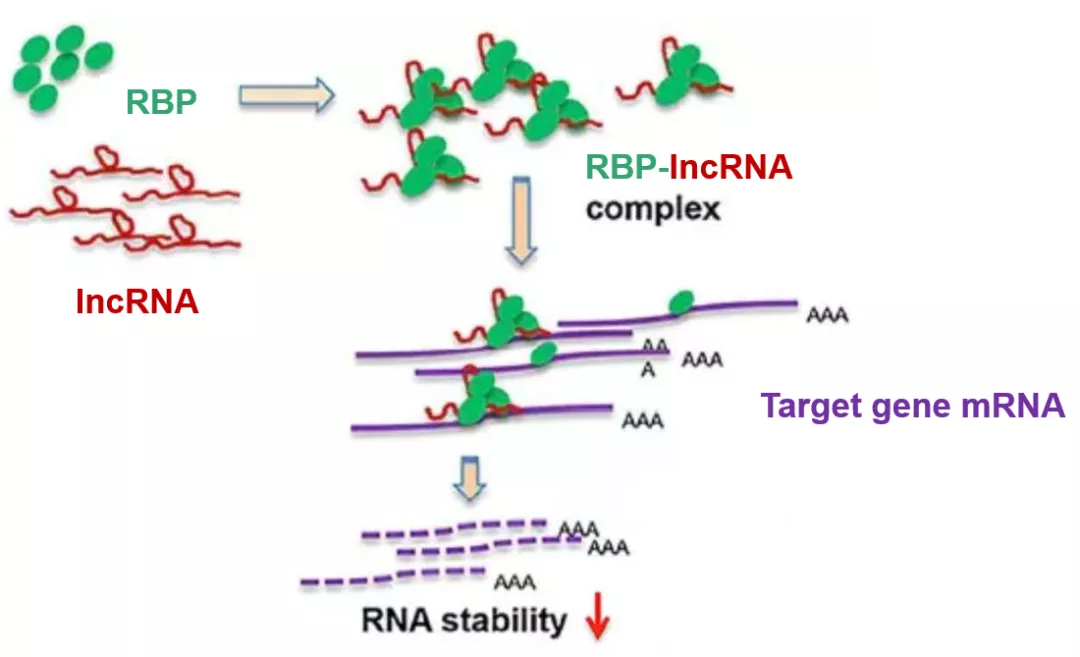
作者一上來就直奔主題,篩選和驗證能夠與lncRNA MEG3結合的RBP:通過有義鏈(sense)和作為陰性對照的反義鏈(anti-sense)的MEG3探針(Fig. 1A),進行RNA pull down后在SDS-PAGE膠上進行考馬斯亮藍染色(非特異的染蛋白質),通過對比anti-sense和sense鏈的差異條帶(Fig. 1B,箭頭所示),將該部分膠進行質譜分析,發現其中所包含的幾十種蛋白中含有PTBP1(正文未展示,補充材料),接著再通過RNA pull down(Fig. 1C)以及RIP(Fig. 1D)實驗進一步確認了MEG3與PTBP1的結合。
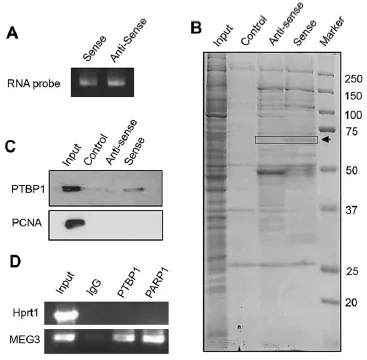

接下來,作者通過預測軟件(RNAmap,starbase 2.0)找到了與PTBP1結合的mRNA SHP(Fig. 2A)。然后在三種細胞中通過RIP驗證了SHP mRNA與PTBP1的結合,并且這種結合會受到MEG3過表達的促進(Fig. 2B)。發現沒有,與ceRNA的競爭結合的機制相反,lncRNA在結合RBP后,并不會導致與靶基因mRNA結合的RBP減少,而是促進RBP與mRNA的結合。進一步地,通過RNA pull down實驗,作者驗證了PTBP1與SHP mRNA的哪一部分結合(CDS,3’UTR)(Fig. 2C)。最后,通過mRNA降解實驗,作者驗證了過表達MEG3和(或)PTBP1能夠促進SHP mRNA的降解(Fig. 2D)。
至此,我們可以跟著下面所示的這篇文獻的思路圖,總結lncRNA/RBP/靶分子的研究套路,先通過RNA pull down/質譜篩選和確認與lncRNA結合的RBP分子,并使用RIP和RNA pull down進行驗證;再通過預測找到與RBP結合下游靶分子mRNA,通過RIP,RNA pull down驗證lncRNA是否影響RBP與mRNA的結合,最后通過mRNA降解實驗驗證lncRNA以及RBP對mRNA的表達的變化的影響。
小拓展1:RBP分子并不像miRNA或是EZH2一樣,僅對下游mRNA或基因產生抑制。在上文分享的例子中,PTBP1是降解靶基因的mRNA的,但是也有文獻報道PTBP1能夠增加某些mRNA的穩定性(Long noncoding RNA ANCR inhibits the differentiation of mesenchymal stem cells toward definitive endoderm by facilitating the association of PTBP1 with ID2),或通過選擇性剪切的方式調控mRNA(PTBP1-Mediated Alternative Splicing Regulates the Inflammatory Secretome and the Pro-tumorigenic Effects of Senescent Cells),這也使得RBP的設計套路更加靈活。
小拓展2:關于lncRNA到底是如何影響RBP與mRNA的結合,與上面的EZH2一樣,一般也少有深入研究,誠如上述這篇14分的hepatology也只是在討論里闡述了幾種可能的機制(見下方截圖),留下一句future studies would be necessary讓人浮想聯翩,想要深入探索的同學不妨朝這方面入手(慎重),想要在自己的討論里吹吹水的同學也可以多作借鑒(推薦)。
關于lncRNA與RBP的技術總結
1. 關鍵的驗證內容是lncRNA與RBP的結合,以及lncRNA對RBP與mRNA結合穩定性的調控
2. 一般來說不需要再深入研究lncRNA與RBP的機制
3. RBP存在功能多樣性,可以正向,負向調控下游mRNA的穩定性
4. 如果沒有做質譜的條件,也可以參考預測軟件的結果找到lncRNA可能結合的RBP
好了,考慮到大家可能已經開始出現看文獻時易發的頭暈,惡心的癥狀,本期先為大家介紹兩種lncRNA在離開miRNA后的機制研究方向:第一種是lncRNA通過調控H3K27me3組蛋白甲基化;第二種是lncRNA通過RBP調控mRNA的穩定性,覺得有點啟發的同學可以在清醒后繼續關注,在下一期中我們會為大家帶來另外兩種不同于ceRNA的機制。


電話:400-691-6686
在線時間:8:00-24:00

微信關注公眾號

 湘公網安備
湘公網安備